1. Knothe, H., et al., Transferable resistance to cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of Klebsiella pneumoniae and Serratia marcescens. Infection, 1983. 11(6): 315-7.
2. Barthélémy, M., et al., Single amino acid substitution between SHV-1 β-lactamase and cefotaxime-hydrolyzing SHV-2 enzyme. FEBS Lett, 1988. 231(1): 217-20.
3. Bush, K. and G.A. Jacoby, Updated functional classification of β-lactamases. Antimicrob Agents Chemother, 2010. 54(3): 969-76.
4. Paterson, D.L. and R.A. Bonomo, Extended-spectrum β-lactamases: a clinical update. Clin Microbiol Rev, 2005. 18(4): 657-86.
5. Nordmann, P., M.F. Lartigue, and L. Poirel, β-lactam induction of ISEcp1B-mediated mobilization of the naturally occurring bla(CTX-M) β-lactamase gene of Kluyvera
ascorbata. FEMS Microbiol Lett, 2008. 288(2): 247-9.
6. Poirel, L., P. Kampfer, and P. Nordmann, Chromosome-encoded Ambler class A β-lactamase of Kluyvera georgiana, a probable progenitor of a subgroup of CTX-M extendedspectrumβ-lactamases. Antimicrob Agents Chemother, 2002. 46(12): 4038-40.
7. Matsumoto, Y., et al., Novel plasmid-mediated β-lactamase from Escherichia coli that inactivates oxyimino-cephalosporins. Antimicrob Agents Chemother, 1988. 32(8): 1243-6.
8. Bauernfeind, A., H. Grimm, and S. Schweighart, A new plasmidic cefotaximase in a clinical isolate of Escherichia coli. Infection, 1990. 18(5): 294-8.
9. Barthélémy, M., et al., Close amino acid sequence relationship between the new plasmid-mediated extended-spectrum β-lactamase MEN-1 and chromosomally encoded
enzymes of Klebsiella oxytoca. Biochim Biophys Acta, 1992. 1122(1): 15-22.
10. Ishii, Y., et al., Cloning and sequence of the gene encoding a cefotaxime-hydrolyzing class A β-lactamase isolated from Escherichia coli. Antimicrob Agents Chemother, 1995.39(10): 2269-75.
11. Bauernfeind, A., et al., Sequences of β-lactamase genes encoding CTX-M-1 (MEN-1) and CTX-M-2 and relationship of their amino acid sequences with those of otherβ
-lactamases. Antimicrob Agents Chemother, 1996. 40(2): 509-13.
12. Livermore, D.M., et al., CTX-M: changing the face of ESBLs in Europe. J Antimicrob Chemother, 2007. 59(2): 165-74.
13. Livermore, D.M. and P.M. Hawkey, CTX-M: changing the face of ESBLs in the UK. J Antimicrob Chemother, 2005. 56(3): 451-4.
14. Bonnet, R., et al., Effect of D240G substitution in a novel ESBL CTX-M-27. J Antimicrob Chemother, 2003. 52(1): 29-35.
15. Karim, A., et al., Plasmid-mediated extended-spectrum β-lactamase (CTX-M-3 like) from India and gene association with insertion sequence ISEcp1. FEMS Microbiol Lett, 2001. 201(2): 237-41.
16. Novais, A., et al., Evolutionary trajectories of β-lactamase CTX-M-1 cluster enzymes: predicting antibiotic resistance. PLoS Pathog, 2010. 6(1): e1000735.
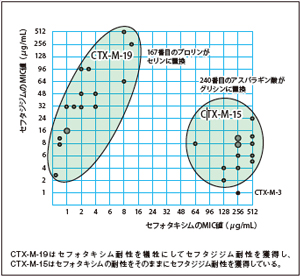
17. Kimura, S., et al., Role of a mutation at position 167 of CTX-M-19 in ceftazidime hydrolysis. Antimicrob Agents Chemother, 2004. 48(5): 1454-60.
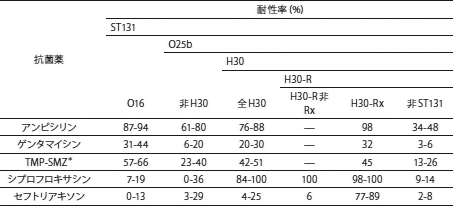
18. Banerjee, R. and J.R. Johnson, A new clone sweeps clean: the enigmatic emergence of Escherichia coli sequence type 131. Antimicrob Agents Chemother, 2014. 58(9): 4997-5004.
19. Nicolas-Chanoine, M.H., X. Bertrand, and J.Y. Madec, Escherichia coli ST131, an intriguing clonal group. Clin Microbiol Rev, 2014. 27(3): 543-74.
20. Price, L.B., et al., The epidemic of extended-spectrum β-lactamase producing Escherichia coli ST131 is driven by a single highly pathogenic subclone, H30-Rx. MBio, 2013. 4(6):e00377-13.
21. Olesen, B., et al., Temporal trends in antimicrobial resistance and virulence-associated traits within the Escherichia coli sequence type 131 clonal group and its H30 and H30-Rx subclones, 1968 to 2012. Antimicrob Agents Chemother, 2014. 58(11): 6886-95.
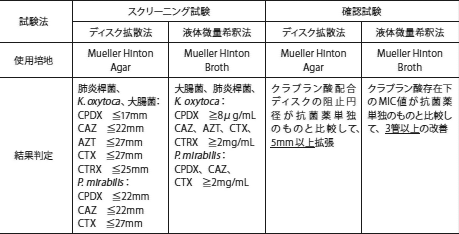
22. Clinical and Laboratory Standards Institute, Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. M100-S25. Vol. 35. 2015,
Wayne, Pennsylvania, USA: CLSI.
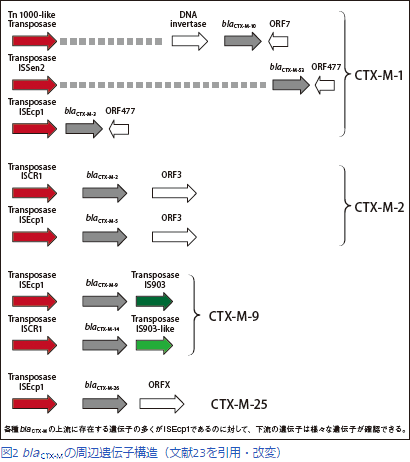
23. Canton, R., J.M. Gonzalez-Alba, and J.C. Galan, CTX-M Enzymes: Origin and Diffusion. Front Microbiol, 2012. 3: 110.
24. Trecarichi, E.M., R. Cauda, and M. Tumbarello, Detecting risk and predicting patient mortality in patients with extended-spectrum β-lactamase producing Enterobacteriaceae bloodstream infections. Future Microbiol, 2012. 7(10): 1173-89.
25. Woerther, P.L., et al., Trends in human fecal carriage of extended-spectrum β-lactamases in the community: toward the globalization of CTX-M. Clin Microbiol Rev, 2013.26(4):744-58.
26. Ewers, C., et al., Extended-spectrum β-lactamase producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin Microbiol Infect, 2012. 18(7): 646-55.
27. Lazarus, B., et al., Do human extraintestinal Escherichia coli infections resistant to expanded-spectrum cephalosporins originate from food-producing animals? A systematic review. Clin Infect Dis, 2015. 60(3): 439-52.
28. Michael, G.B., et al., Emerging issues in antimicrobial resistance of bacteria from food-producing animals. Future Microbiol, 2015. 10: 427-43.
29. Dubois, V., et al., CTX-M-producing Escherichia coli in a maternity ward: a likely community importation and evidence of mother-to-neonate transmission. J Antimicrob Chemother, 2010. 65(7): 1368-71.
30. Eppes, C.S. and S.L. Clark, Extended-spectrum β-lactamase infections during pregnancy: a growing threat. Am J Obstet Gynecol, 2015. [Epub ahead of print] doi: 10.1016/j.ajog.2015.03.020.
31. Kristóf, K., et al., Extended-spectrum β-lactamase producing Klebsiella spp.